Новости
ООО "Компания Новые технологии" - ТОП 1000 Поставщиков
ООО "КНТ" вошла в число успешных российских поставщиков по итогам работы на площадке B2B-Center в 2025 году.
ТОП поставщиков составлен по итогам работы компаний на площадке B2B-Center в 2025 году.
Исследование проводилось среди 569 133 организаций, зарегистрированных в системе.

Представители компании в регионах
Руководитель:
Рыжов Михаил Викторович
8 909 239 00 88
Коммерческий директор:
Прокопов Дмитрий
Александрович
8 905 158 75 00
Директор по продажам
Талащенко Антон
Евгеньевич
8 910 736 01 23
Белгородский регион
Тарасиков Виталий (КРС)
8 960 640 13 99
Липецкий регион
Москвитин Игорь
8 903 877 40 06
Воронежский регион
Свешникова Екатерина
8 950 870 40 77
Брянский регион
Михайлов Юрий
8 960 675 94 00
Орловский регион
Поздняков Михаил
8 920 261 36 94
Курский регион
Михайлов Юрий
8 960 675 94 00
Рыжов Михаил
8 909 239 00 88
Рязанский регион
Москвитин Игорь
8 903 877 40 06
Калужский и Тульский регионы
Поздняков Михаил
8 920 261 36 94
Ленинградский регион
Армоник Эвальд
8 903 872 80 79
Технический специалист
по кормлению КРС
Абдулин Алексей 8 951 332 03 31
Специалист по кормлению ООО "ТехКорм"
Новиков Сергей 8 910 466 72 20
Независимый консультант по свиноводству и птицеводству
Василаки Игорь Андреевич 8 980 370 15 15
Аминокислотное питание коров: влияние на воспроизводство и качество потомства. Часть 2

Перед вами вторая статья из серии «Аминокислотное питание коров», посвящённой влиянию лимитирующих аминокислот, метионина и лизина, на эффективность молочного животноводства.
В предыдущей статье (Д. Г лухов, 2018, Аминокислотное питание коров: влияние на здоровье животных) были рассмотрены функции этих аминокислот, затрагивающие различные аспекты здоровья животных: регуляция жирового и глюкозного обмена, защита от окислительного стресса, снижение интенсивности воспалительных процессов. И хотя здоровье коров наряду с молочной продуктивностью зачастую находятся под более пристальным вниманием собственников и управляющих молочных ферм, так как они напрямую влияют на прибыль, не стоит также забывать о показателях воспроизводства, поскольку именно они гарантируют рентабельность предприятия в долгосрочной перспективе.
Данные за последние десятилетия свидетельствуют о неутешительной тенденции снижения показателей воспроизводства по мере роста молочной продуктивности. Так, например, в период с 1970 по 2000 год межотёльный интервал в среднем увеличился с 13,5 до 14,7 месяцев, а индекс осеменения вырос с 1,8 до 3,0 (29).
Снижение уровня воспроизводства или яловость являются следствием множества причин. Наряду с некорректным управлением стадом, проблемами с обменом веществ, или внешними факторами (14, 42), неправильное кормление также существенно влияет на репродуктивные функции. Защищённые от распада в рубце метионин и лизин в некоторых случаях способны улучшить показатели воспроизводства как путём воздействия на эффективность использования азота корма, так и в качестве биологически активных веществ, стабилизирующих течение беременности и улучшающих качество потомства.
Влияние эффективности использования азота на воспроизводство.
Для стимуляции и поддержания высокой молочной продуктивности, особенно в начале лактации, коровам зачастую скармливаются рационы с высоким содержанием сырого протеина (>17%), однако это может негативно сказываться на репродуктивной функции (8), так как аминокислоты, которые не были использованы в процессах синтеза белка, расщепляются до аммиака. Поскольку последний токсичен для организма, он обезвреживается путём преобразования в мочевину и выводится с мочой. Однако синтез мочевины - это энергозатратный процесс, соответственно скармливание избыточного количества белка приводит к дополнительным затратам энергии, которая могла бы быть использована, например, на восстановление организма после отёла. Кроме того, мочевина сама по себе может снижать репродуктивную функцию (21).
С этой точки зрения хорошими индикаторами эффективности кормления и воспроизводства являются такие показатели как концентрация азота мочевины в плазме крови (АМП) и в цельной крови (АМК). Эти показатели тесно связаны друг с другом и могут быть вычислены один из другого по следующей формуле:
АМП, мг/100 мл = (1.021 X АМК) + 0.399
Ещё одним хорошим индикатором эффективности белкового обмена в организме коров служит азот мочевины молока (АММ) (39). Данный показатель может использоваться для оценки правильности кормления в силу того, что он напрямую коррелирует с азотом мочевины крови (АМК), так как мочевина легко переходит из крови в другие ткани (31). Их взаимосвязь можно выразить следующей формулой:
АММ, мг/100 мл = (0.620 X АМК) + 4.75
Целевое значение концентрации АММ составляет 8-10 мг/100 мл (44), что соответствует 171-214 мг/л мочевины.
Отрицательное влияние повышенных концентраций азота мочевины в плазме, цельной крови и молоке на воспроизводство было доказано во множестве исследований. Melendez с соавторами в 2003 году обобщили часть этих данных, результат представлен в таблице 1.
Таблица 1— Влияние концентрации азота мочевины плазмы крови, цельной крови и молока (АМП, АМК и АММ соответственно) на показатели воспроизводства у молочных коров (32)
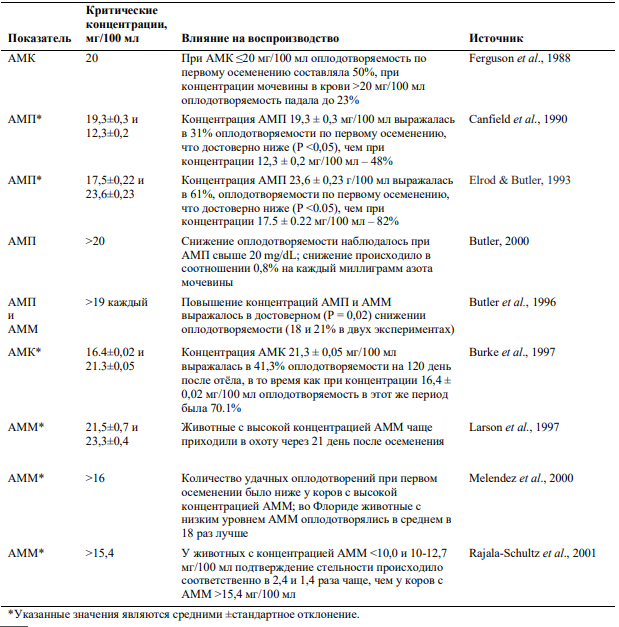
Снижение оплодотворяемости может происходить из-за того, что высокие концентрации азота мочевины в плазме крови влияют на pH среды в матке (15, 16, 38) и концентрацию мочевины, магния, калия фосфора и цинка (26). Слишком сильные изменения среды внутри матки, особенно на ранних этапах развития эмбриона, могут существенно повлиять на его выживаемость. Помимо этого, высокий уровень АМП может влиять и на качество эмбрионов. Например, в исследовании, проведённом на 180 лактирующих коровах, простимулированных на суперовуляцию (лекарственная или гормональная стимуляция на одновременную выработку большого количества фолликулов, один из этапов технологии трансплантации эмбрионов), было выявлено, что от животных с уровнем азота мочевины в молоке от 12 до 18 мг/100 мл было получено большее число пригодных к трансплантации эмбрионов, чем от животных с концентрацией АММ выше 18 мг/100 мл (7,4 ± 0,6 против 4,6 ± 0,6; P < 0,05) (28). Также было установлено, что повышенные уровни азота мочевины в плазме крови снижали жизнеспособность эмбрионов овец и тормозили их развитие и in vivo, и in vitro (4, 30).
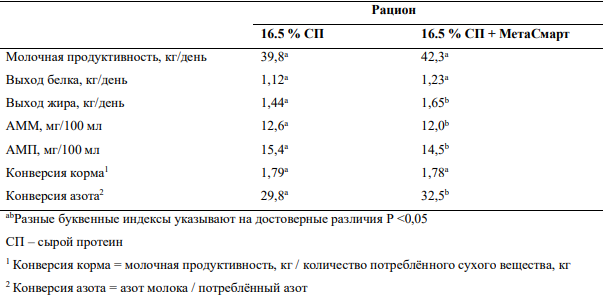
Введение в рационы жвачных лимитирующих аминокислот, метионина и лизина, может существенно улучшить эффективность использования азота, что не только положительно повлияет на продуктивность, но и снизит концентрации азота мочевины в плазме крови и, соответственно, в молоке (Таблица 2).
Таблица 2— Влияние изопропилового эфира 2-гидрокси-4-(метилтио) бутановой кислоты (HMBi) на молочную продуктивность и концентрации азота мочевины в плазме крови и молоке (АМП и АММ соответственно) (43)
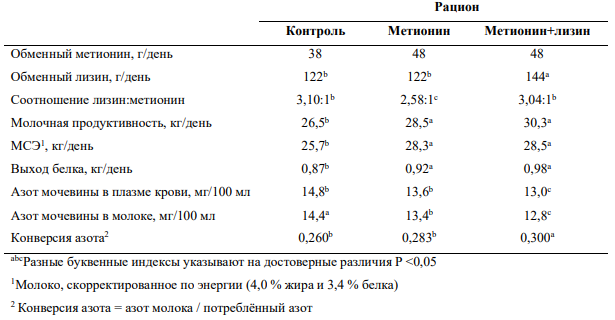
Более того, введение сразу обеих лимитирующих аминокислот в правильном соотношении может повлиять на снижение АМП м АММ сильнее, чем использования каждой из них по отдельности. Так, например, эксперимент Wang и соавторов (2010) показал, что одновременное введение метионина и лизина в рацион, содержащий 16,5% сырого протеина, оказывало больший эффект на молочную продуктивность и концентрации азота мочевины в плазме крови и молоке, чем введение одного метионина (Таблица 3).
Таблица 3— Влияние введения в рацион метионина и лизина на молочную продуктивность, эффективность использования азота и концентрации азота мочевины в плазме крови и молоке (45)
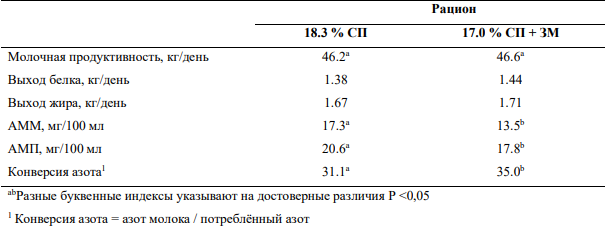
Введение защищённых аминокислот позволяет также снизить количество сырого протеина в рационе без потери молочной продуктивности, что повлечёт ещё более существенное снижение концентрации азота мочевины в крови и молоке (Таблица 4).
Таблица 4— Влияние включения в рацион защищённого метионина (ЗМ) наряду со снижением количества сырого протеина (СП) на молочную продуктивность и концентрации азота мочевины в плазме крови и молоке (АМП и АММ соответственно) (33)
Исходя из всего вышесказанного можно предположить, что балансирование рационов по аминокислотам может положительно повлиять на воспроизводство как минимум за счёт снижения концентрации азота мочевины в крови и, как следствие, во внутриматочной среде.
Влияние отдельных аминокислот на воспроизводство
Как уже упоминалось выше, аминокислоты могут оказывать положительное влияние на физиологические процессы независимо от их участия в синтезе белков. Это можно назвать «функциональными эффектами» аминокислот. Наиболее изученными с точки зрения влияния на воспроизводство «функциональными аминокислотами» являются метионин и аргинин (36, 47).
Для того, чтобы оценить насколько велико значение аминокислот в процессе размножения, давайте для начала кратко рассмотрим, что происходит на начальных этапах развития эмбриона.
Итак, оплодотворение яйцеклетки и первые стадии развития зародыша, называемые дроблением, происходят в яйцеводе. Первоначально питание эмбиона происходит лишь за счет запасов желтка, накопленных в яйцеклетке, но уже на поздних стадиях дробления зародыш начинает использовать питательные вещества, получаемые из содержимого секрета, выделенного железистыми клетками яйцевода и матки. Примерно к 6-7 дню после овуляции эмбрион развивается до стадии бластоцисты и достигает рога матки. Примерно на 9 день зародыш сбрасывает блестящую оболочку и где-то на 17 день прикрепляется к стенке матки. На 25-28 сутки становится возможным питание эмбриона через формирующуюся плаценту. Всё время до этого зародыш питается за счёт желтка и выделений маточных желёз, т.е. его жизнеспособность и степень обеспеченности необходимыми для развития питательными веществами, в том числе аминокислотами, во многом зависят от внутренней среды матки. Таким образом очень важно понимать, как могут изменяться концентрации тех или иных веществ, в частности аминокислот, в матке в процессе развития эмбриона (11).
Hugentobler и соавторы (2007) обобщили данные по концентрации аминокислот в плазме крови (измерение на 0, 2, 3, 4 и 6 день эстрального цикла), в яйцеводах и матке (измерение на 6, 8 и 14 день эстрального цикла) гибридных нетелей мясного направления. Изменений концентрации аминокислот в зависимости от стадии эстрального цикла не наблюдалось. Однако концентрации 9 из 20 аминокислот в яйцеводах были существенно выше, чем в плазме крови, что свидетельствует о в клетках яйцеводов наличии механизмов, позволяющих увеличивать концентрацию аминокислот в своих выделениях для обеспечения ими зародыша. В матке также наблюдались повышенные концентрации многих аминокислот по сравнению с плазмой крови. Наибольшей концентрации в яйцеводах и матке достигали аспарагиновая кислота, аспарагин и глутаминовая кислота.
Помимо способности постоянно поддерживать более высокою по сравнению с кровью концентрацию аминокислот, клетки яйцеводов и матки могут дополнительно увеличивать концентрации некоторых аминокислот в своих выделениях в период активного роста эмбриона (так называемая элонгация, происходит на 14-18 день). Было проведено три исследования по определению концентраций аминокислот яйцеводах и матке в этот период, два на овцах (18) и одно на коровах (20). Исследования Gao и соавторов (2009) дополнительно подтвердили, что у небеременных животных на 10-16 день цикла концентрации аминокислот в секрете железистых клеток матки не изменялись, тогда как у суягных овец концентрации отдельных аминокислот возрастали 3-23 раза.
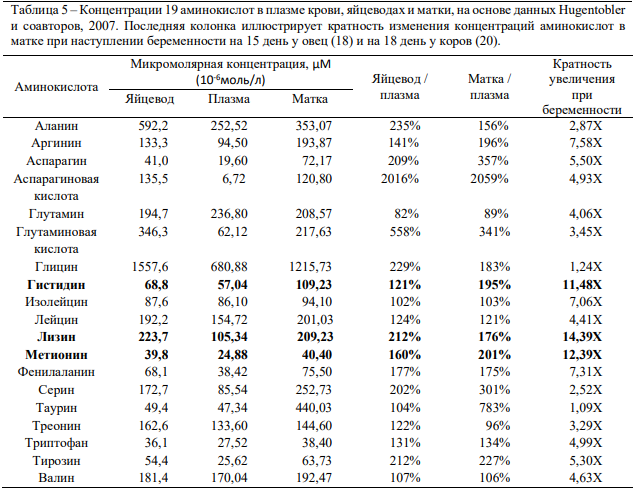
Wiltbank и соавторы (2014) суммировали данные описанных выше исследований (18, 20, 22), чтобы наглядно проиллюстрировать существенные различия концентраций аминокислот в плазме крови, яйцеводах и матке, а также их многократное возрастание в период активного роста эмбриона. Полученные результаты, позволяющие получить представление об изменениях количества аминокислот в матке на ранних стадиях беременности, представлены в таблице 5.
Интересно то, что при беременности в матке сильнее всего возрастала концентрация тех аминокислот, которые признаны лимитирующими молочную продуктивность у жвачных, то есть метионина, лизина и гистидина (более чем десятикратное увеличение во всех трёх исследованиях). Соответственно можно предположить, что недостаточное поступление в организм матери этих аминокислот в период, как минимум, элонгации эмбриона, может негативно сказаться на темпах его роста или развитии плаценты (46).
Таблица 5 - Концентрации 19 аминокислот в плазме крови, яйцеводах и матки, на основе данных Hugentobler и соавторов, 2007. Последняя колонка иллюстрирует кратность изменения концентраций аминокислот в матке при наступлении беременности на 15 день у овец (18) и на 18 день у коров (20).
Как уже упоминалось выше, наиболее изученными аминокислотами с точки зрения воздействия на воспроизводство у животных являются аргинин и метионин. При этом исследований на жвачных в этой области достаточно мало. На сегодняшний день относительно большой объём данных о влиянии на репродуктивную функцию и развитие эмбриона у коров есть только для метионина.
Здесь стоит упомянуть один из наиболее ранних, но достаточно интересных опытов, проведённых Coelho и соавторами в 1989 году. Исследователи использовали сыворотку крови лактирующих коров для выращивания крысиных эмбрионов на стадии органогенеза (9,5 дней после зачатия), которые нормально развиваются в плазме крови крыс без необходимости введения каких-либо дополнительных питательных веществ. В силу этого их часто используют для определения потребностей зародыша в конкретных веществах в процессе развития или выявления токсичных для эмбрионов веществ в крови животных разных видов (13). В ходе эксперимента выяснилось, что при помещении эмбрионов в чистую сыворотку коровьей крови, их развитие происходило с нарушениями (недозакрытие нервной трубки, не развивались глаза и жаберные дуги развивались эмбрионы неправильной формы). При добавлении в сыворотку комплекса аминокислот и витаминов развитие шло нормально. Введение только аминокислот без витаминов также позволяло зародышам развиваться без патологий. Использование комплекса аминокислот без метионина снова привело к появлению нарушений, а вот введение только метионина снова нормализовало развитие эмбрионов. После этого было решено провести дополнительный эксперимент, в ходе которого для выращивания зародышей была использована сыворотка крови коров, дополнительно получавших защищённый метионин (110 г/день). В ней развитие эмбрионов также шло без патологий. Одним из выводов исследования было предположение о том, что концентрация метионина в крови коров столь низкая, что его не хватает даже для обеспечения потребностей крысиных зародышей, а правильное развитие собственных эмбрионов происходит, вероятно, только при наличии механизмов, позволяющих накапливать необходимые питательные вещества.
Ещё один интересный эксперимент по установлению необходимости метионина для нормального развития эмбрионов коров проводилось Ikeda и соавторами в 2012 году. Исследователи вводили в эмбриональные культуры КРС дополнительный метионин, либо этионин. Последний является антагонистом метионина и блокирует его метаболизм в тканях. Этионин не влиял на развитие зародышей в стадии морулы (3-5 день развития), но практически полностью блокировал развитие морулы в бластоцисту (Контроль = 38,5%, Этионин = 1,5%). Соответственно метионин играет важную роль уже на столь ранней стадии эмбриогенеза - без него зародыш просто не будет развиваться дальше.
Однако метионин может отвечать не только за нормальное развитие эмбриона, но и за накопление питательных веществ, необходимых для поддержания жизни развивающегося организма. Недавнее исследование Acosta и соавторов (2016) показало, что введение в рацион молочных коров защищённого метионина на последних стадиях развития фолликула и ранних этапах развития эмбрионов (до 7 дня) приводило к накоплению большего количества жира в зародышах. Таким образом, метионин влияет на выживаемость эмбриона, как минимум до момента сброса блестящей оболочки, пока он питается за счёт запасов желтка, а возможно и до момента его прикрепления к стенке матки и развития плаценты, поскольку именно накопленный жир будет являться одним из основных источников энергии для зародыша. Подтверждением этому может служить более низкий уровень ранней эмбриональной смертности у коров второй и более лактации, дополнительно получавших защищённый метионин (11).
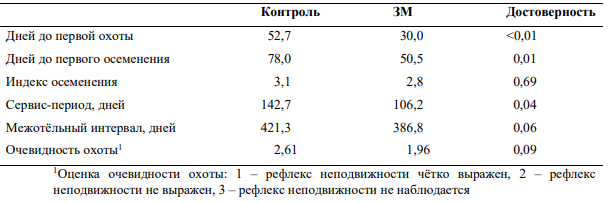
На ближнем востоке также были получены данные, подтверждающие положительное влияние защищённого метионина на показатели воспроизводства. Ardalan и соавторы (2010) отметили существенное снижение сервис-периода и индекса осеменения (р < 0,05) у коров голштинской породы, получавших 18 г защищённого от распада в рубце метионина в день в течение 4 недель до и 20 недель после отёла (Таблица 6) . Также была отмечена тенденция к уменьшению дней до первой охоты. Интересно, что введение 60 г защищённого холина в дополнение к метионину оказывало существенно больший эффект на воспроизводство (Таблица 6).
Таблица 6 — Влияние защищённого метионина (ЗМ) и защищённого холина на показатели воспроизводства у коров голштинской породы (3)

Nikkhah и соавторы (2013) также оценивали влияние защищённого метионина на воспроизводство. У коров, получавших метионин, наблюдалось уменьшение количества дней до первой охоты, снижение индекса осеменения, уменьшение сервис-периода и межотёльного интервала (Таблица 7).
Таблица 7 — Влияние защищённого метионина (ЗМ) на показатели воспроизводства у коров голштинской породы в Иране в летний период (34).
Улучшение качества потомства: эпигенетические эффекты метионина
Прежде чем рассмотреть эту тему необходимо кратко пояснить, что же такое эпигенетика. Она представляет собой изучение закономерностей эпигенетического наследования - изменения экспрессии генов или фенотипа клетки, вызванных механизмами, не затрагивающими последовательности ДНК. Проще говоря, это раздел генетики, изучающий факторы и механизмы, отвечающие за «включение» и «выключение» определённых генов на определённых этапах развития организма. Наиболее изученным к настоящему моменту эпигенетическим механизмом является метилирование ДНК, в котором метионин играет ключевую роль. Таким образом обеспечение необходимого уровня метионина в организме животных на определённой стадии беременности может благоприятно повлиять н только на её течение, но и улучшить физиологические показатели потомства. Например, в опыте, проведённом Sinclair и соавторами (2007), овцам до осеменения и в течение 6 дней после него скармливался рацион, не удовлетворяющий потребности в метионине. Затем полученные от этих животных эмбрионы, чьё развитие было оценено как нормальное, трансплантировались контрольными животным, не испытывавшим недостатка в метионине. После окота проводилась оценка ягнят. Животные, полученные от матерей, испытывавших недостаток в метионине, имели существенные отличия в кровяном давлении и иммунной функции. Penagaricano и соавторы (2013) провели схожее исследование на коровах. Они проверяли эмбрионы, полученные от коров, получавших и не получавших метионин с рационом, на различия в экспрессии генов (реализации генетического материала). Оценивались только эмбрионы высшего качества, полученные от определённых коров, использовались методы, позволяющие отследить экспрессию любых генов. Примечательно, что даже небольшая разница в концентрации метионина в крови приводила к существенным различиям в экспрессии генов в эмбрионах. Всего было происследовано 10662 гена от коров обеих групп. Статистически достоверные различия были обнаружены в экспрессии 276 генов. В частности, в контрольной группе наблюдалась более сильная экспрессия 200 генов, в опытной - 76 генов, то есть большая часть этих генов была «выключена» у животных, дополнительно получавших метионин. Вполне возможно, что подавление экспрессии этих генов происходит целенаправленно до достижения определённого этапа развития, поскольку многие из них вовлечены в регуляцию иммунной функции или отвечают за развитие на более поздних стадиях и их преждевременное «включение» может быть критичным как для дальнейшего протекания беременности так и для нормальной работы иммунной системы телёнка после рождения.
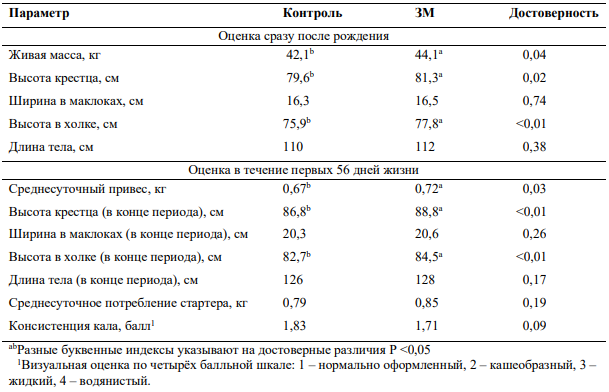
Таким образом, обеспечение потребностей коров в метионине может повлиять на экспрессию генов развивающегося эмбриона, что в свою очередь может положительно сказаться на течении беременности и качестве потомства. Это подтверждают результаты совсем недавнего исследования, которые были опубликованы осенью 2018 года (2). В данном опыте сравнивали телят, полученных от двух групп коров третьей лактации, которым в течение сухостойного периода скармливали одинаковые по составу кормов и питательности рационы. Рацион раннего сухостоя (45-29 день до отёла) содержал 5,57 МДж чистой энергии лактации (ЧЭЛ) и 13,9% сырого протеина (СП), позднего сухостоя (28 дней до отёла) - 6,16 МДж ЧЭЛ и 15,3% СП. Различие заключалось лишь в том, что в рацион опытной группы в течение последних 28 дней стельности был дополнительно включён защищённый метионин, с тем расчётом, чтобы соотношение обменного метионина к обменному лизину составляло 2,81:1. При рождении телят взвешивали, а также измеряли длину тела, высоту в холке, высоту крестца, ширину зада в маклоках. Всем новорожденным животным выпаивали молозиво по одной и той же схеме и далее все телята получали одинаковый по составу заменитель молока и стартерный комбикорм. Схемы выпойки и кормления, а также условия содержания для всех животных были одинаковыми. В течение первых 56 дней жизни телят ежедневно фиксировали потребление ими комбикорма и еженедельно оценивали темпы роста по следующим показателям: живая масса, высота в холке, высота крестца, ширина зада в маклоках. Помимо этого, также оценивали консистенцию кала.
Уже при оценке телят сразу после рождения были выявлены достоверные различия по ряду показателей. В частности, телята, рождённые от коров, получавших защищённый метионин, имели большую живую массу, высоту в холке и крестце. В дальнейшем основные отличия были отмечены по следующим параметрам: среднесуточный привес, высота в холке и высота крестца. Кроме того, у потомства животных, получавших метионин, была отмечена тенденция к увеличению потребления стартерного комбикорма и лучшей консистенции кала, то есть телята, полученные от коров опытной группы не только были изначально крупнее, но и быстрее росли и развивались в первые, наиболее важные, 8 недель жизни, а также, вероятно, имели меньше проблем с пищеварением или были более устойчивы к бактериальным инфекциям. Обобщённые результаты опыта представлены в таблице 8.
Исходя из этих данных можно сделать вывод, что скармливание коровам защищённого метионина даже в течение достаточно короткого периода перед отёлом, последних четырёх недель стельности, способно положительно повлиять на темпы роста и развития, а также, вероятно, на иммунную функцию телят в наиболее важный период - в первые 2 месяца жизни, когда идёт активное развитие всех органов, включая молочную железу (5).
Таблица 8 — Влияние включения защищённого метионина (ЗМ) в рационы коров голштинской породы за 28 дней до отёла на параметры роста и развития телят (2).
Заключение
Воспроизводство - это важный аспект, который обеспечит рентабельность молочной фермы в течение долгого времени. Для поддержания высоких показателей воспроизводства необходимо, помимо грамотного управления стадом и работы с генетикой, правильно составлять рационы и отслеживать их эффективность. Это особенно важно в самом начале лактации, когда организм коровы подготавливается к следующему репродуктивному циклу. Особое внимание в этот период должно уделяться контролю концентрации мочевины в молоке, поскольку этот показатель напрямую связан с концентрацией мочевины в крови, а её высокий уровень может существенно ухудшить оплодотворяемость. У животных, получающих правильно составленный рацион, уровень мочевины в молоке должен быть на уровне 170-215 мг. При этом наиболее информативным будет определение мочевины в сборной пробе молока, полученной путём отбора и смешивания проб с каждой дойки в течение дня.
Помимо балансирования по протеину, энергии, витаминам, макро- и микроэлементам, рационы также должны оцениваться по содержанию лимитирующих аминокислот, поскольку последние, будучи многофункциональными веществами, оказывают положительное влияние не только на молочную продуктивность и содержание жира и белка в молоке (12, 19, 23, 35, 41) за счёт повышения эффективности использования азота, но и на ряд важных физиологических процессов в организме коровы как биологически активные вещества и их предшественники. Таким образом, скармливание рационов с невысоким уровнем сырого протеина (<17% на 1 кг сухого вещества) с введением необходимого количества метионина и лизина в правильном соотношении будет наилучшим решением в начале лактации.
Лекция по аминокислотам:
https://m.youtube.com/watch?v=ynQ3D6fcjLM&t=47s
Вебинар по протеиновому питанию жвачных:
https://youtu.be/OOAB0TiTDTY